貝殻成長線分析
貝殻成長線は、貝類の成長にともなって構成された輪文状の形質で、背面では輪文様、切断面では縞模様を呈する。この形質は、生殖活動や疾病などの内的因
子と海水温度、潮
汐、塩分濃度などの外的因子を反映して形成される。つまり、規則的で美しい炭酸カルシウムの結晶を主体とする軟体動物門の貝殻は、生息環境を反映した成長
の履歴を殻として形に残したものと言い
換えられる。
現在行われているこの種の分析では、縞模様の幅と形状の変化を中心として研究が行われ、生物学、水産学、古生物学、考古学で応用が図
られている(ClarkⅡ
1974)。
貝殻成長線分析は、貝殻に残された成長形質の総合的分析であり、単なる「線」の分析に止まらない。研究入門当初は、成長線の本数を数えることに意識が流
れがちであるが、研究が進展するとともに成長形質を総合的に観察する姿勢が必要となることが理解される。成長線と成長線
の間にある成長増量、そしてそれらの集合のありかた、その材質とそれらに影響する上記の内的因子と外的因子までもが総合的に研究されるべきなのである。貝
殻成長
線分析は、そのような成長の痕跡とその発現の因子を追求し、現生の貝殻と遺跡出土の貝殻の比較研究を通して、過去の貝類を解釈することが主眼とするべきで
ある。
なかでも、成長線と成長増量にさまざまな微量元素が固定されており、貝殻成長線分析を踏まえ、これを理化学的に分析することによって生物学的情報を豊富
に得ることが可能である。
この分析の詳細は、富岡(2003)に詳しい。図版や表も、このページより豊富に掲載されているので、興味のある方は参照して頂きたい。
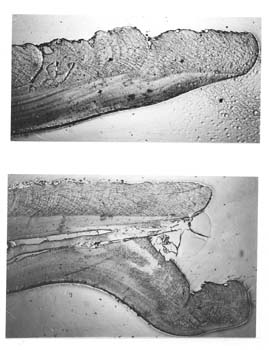
遺跡出土貝類の客観的データと遺物化過程データ
出土した貝類を丁寧に扱って、実証的に考古学的解釈を得るには、さまざまなデータ類の検討が必要になる。最初に貝殻自体の持つ「客観的データ」の検討を
行い、貝類群
集の特徴を把握することが必要である。この段階では、同定に基づいて包含量、サイズ、形態、貝殻成長線分析から導かれる死亡季節や年齢構成のデータ、酸素
同位対比法による古海水温のデータ、貝殻構造のデータが抽出される。
客観的データの最も基礎となるデータは、貝殻の計測データである。貝殻成長線分析は本来は、全ての貝類に実施することが可能であるが、ここでは日本考古
学で一般的な斧足綱について話を絞って紹介を行なう。斧足綱の貝殻成長線のための貝殻切
断ラインと計測点を掲げる。この切断ラインは、貝殻の殻高
の腹縁側の測定点と殻頂を結んだ直線に一致するように設定されている。
さらに貝殻に残されたさまざまな痕跡や母集団の復元を行い、客観的データの資料操作性を検討する遺物化過程のデータ分析が必要になる。具体的には、穿孔
や傷が人為的なものか、肉食の生物によるものかといった加工痕跡の検討や、貝殻が黒色に変化しているのは火を受けた痕跡か、人為的な付着物か、それとも自
然の営為なのかといった観察を行うのである。これらの研究を統合する分析法として、貝殻成長線分析の有効性が指摘できるし、考古学の貝類分析に欠かせない
ものであることが理解できよう。
貝殻成長線分析の原理との解釈
貝殻成長線には、強く太い線と弱く細い線、さらに弱く途切れがちの線がある。潮が大きくひいたり、外敵に襲われたりして強く長い間殻を閉じた時に「強い
線」が形成されると考えられ、軽い干潮等のなんらかの原因で短時間軽く殻
を閉じた時に形成されたものが「弱い線」を形成するものと考えられる。これらの成長線の形成過程の関係は明確に実証されておらず、複数の説が存在してい
る。大ざっぱに分類して、体内時計の様な「内的因子」を強調する説と潮の干満の影響を大きいものとして評価する「外的因子」強調説が挙げられる。
この2説の議論は、考古学に応用する上でも重要なのであるが、いずれの説にしても、内的環境と外的環境の双方の変化を反映して成長線と成長増量が形成さ
れていると考えられることから、ここではその相違を強調することはしない。
「強い線」は線の太さとその形状からAタイプとBタイプ、「弱い
線」は同様にC〜Eタイプに分類される(小池1975)。このタイプについて の研究を簡単に以下にまとめる。小池裕子によるハマグリの放流実験(小池
1975)や東北大考古学研究室と福島県水産試験場松川浦分場でのアサリの放流実験(須藤編
1995)では、A線とB線の合計が放流日数とほぼ一致している。ただし、この線は同日同時間でも生息深度によってA線が生じたりB線が生じたり、あるい
はそれより弱いC以下の線が生じたりする可能性がある。成長線に関するこのような貝類の特性を貝種や生息深度毎に把握することが必要不可欠であり、成長線
計数の際の基礎データとして提示するべきである。C〜Eタイプの成長線はアクセサリーラインやサブディリーライン等と呼ばれることがある。死亡季節推定を
実施する場合、この成長線計数を①A線のみで実施する方法.②A・B線をあわせた状態で実施方法.で微妙な数値の変化が生じる可能性がある。そのため最終
年の冬輪とその一年前の冬輪の間を計数し、一年間の日数である365に近くなる計数法を選択し、合理的に判断して結果を出している。最終的にこの計数方法
で、カウントに利用される成長線を「日周線」と呼ぶことができる。
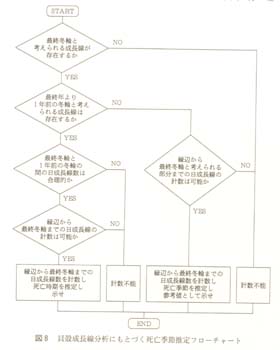
考古学分野での貝殻成長線分析では、貝殻が成長を終了した時期の情報−言い換えれば「死亡時期」−を得ることが主題とされている。「死亡時期」は、−あ
る意味とても奇妙であるのだが−所属年次不詳のまま、以下の作業によって「日付」の形式で算出される。「季節」に変換される場合もある。この「死亡日」
は、「採集日」を反映させている可能性が高いので、貝採集季節の推定に有効と考えられるわけである。
日本で現在行なわれる考古学的貝殻成長線分析のほとんどで、マルスダレガイ科とシジミガイ科が用いられているので、これらの貝類に関する手法を述べる。
まず、腹縁(margin)の保存が良好な資料を選択し、成長線の密な様相である「成長停滞」とその周辺の成長線のあり方を観察する。
成長の停滞は「冬輪」と呼ばれる冬期の成長停滞期が代表的である。それ以外に生殖にともなう成長障害や、夏の成長停滞、外套膜が物理的に傷つけられるア
クシデントによる成長障害が、冬輪にやや似る「成長停滞」「成長障害」を残すが、これらは現生の貝類の研究で成長線パターンが分析されており、冬輪と峻別
されている。
このような成長障害(ディスターバンス)のあり方をパターンとして捉えたものをディスターバンス・パターンと呼ぶ。このディスターバンス・パターンは、
近接した海域のほぼ同じ年齢階梯の同種の貝殻に見られることが指摘されている。第3図は、福島県水産試験場のアサリ生体の放流実験の結果、200m程度離
れた標高の異なる別地点で、類似したディスターバンス・パターンが指摘された例である。
冬輪中心は、日本沿岸部で海水温が最も低下する2月15日と仮定する。そのポイントから、貝殻の腹縁までの日成長線の本数を計数し、2月15日にたす
と、「死亡日」さらに「死亡季節」が算出されるわけである。
「死亡季節」以外にも、母集団の「年齢構成」「成長パターン」等が把握されるが、なかでも重要なデータは「死亡季節」のデータである。貝殻のブロックが
検出された場合、その個体群の「死亡季節」がどのような分布を示すのかは、考古学的解釈に欠かせない情報となるし、古海水温のデータも貝殻成長線分析に
よって把握された季節毎に提示されることが望ましい。また、食用にされた季節と、その処理方法の傾向の把握も季節毎に提示されることが望ましいといえる。
このように、貝類の考古学的解釈に貝殻成長線分析は重要な位置を占めるが、これらの研究を総合しても、遺物化の経路をすべて実証するのは困難である。これ
らを全て明らかにすることよりも、むしろその資料がどれほどの情報を有し、その解釈にはどのような限界が存在しているのか推定する「資料批判」こそが形成
過程分析においては意義が大きいものといえよう。
貝類の比較データ
「客観的データ」と「遺物化過程データ」を、正確に解釈するために必要なデータが「比較データ」として位置づけられる。これにより、前二者のデータが解
釈に導かれる。
低湿地遺跡で貝層が検出された場合、自然貝層か人為的貝層であるのか、問題になることが多い。この際、貝層中に人工遺物が共伴していれば、貝塚の可能性
を尊重して対応することが望ましい。実際にこのような遺跡を発掘・分析すると、自然貝層と考えられるような単調で稚貝を多く含む貝層中に、微細な人工遺物
や、カット・マークを残す哺乳類の骨片などの人為的に廃棄された動物遺存体が検出されることがある。微小魚骨をはじめとする食料残滓を発掘中に検証するこ
とは困難であるから、検出当初より、慎重に発掘に取り組むことが必要なのである。貝層を採集した後の丁寧な分析によって、自然貝層か人工的な貝層か、ある
いはその両方の面を持っているのか解釈するべきであろう。自然貝層と人為的廃棄層を含んでいる場合、現在われわれが海浜で見かける自然死の貝殻と調理の廃
棄物である貝殻やゴミ類が混ざった状態に類似している。解釈にあたっては、貝類群集の稚貝の包含率、貝殻表面の摩耗や風化、肉食貝類などによる摂餌痕跡な
どの「客観的データ」と「遺物化過程データ」とともに、珪藻・花粉などの微化石分析や、土壌pHなどの地質分析を合わせて行い、「環境復元」や「共伴関
係」といった「比較データ」を検討し、総合的に解釈を行う必要性がある。
また、貝殻は加工され装飾品や生活用具として出土することも多い。縄文時代、弥生時代には腕輪や玉、貝殻腹縁を利用した篦や刃器に加工された貝製品が出
土する。このように小破片に加工された貝殻も、切断面や表面等の成長線が顕微鏡観察できれば、科・属・種が同定できる可能性がある。
中近世の遺跡からはイタヤガイやアカニシに柄を取り付けた杓子である「貝杓」(カイジャク)や貝殻製碁石が出土する。これらは、食用貝あるいは非食用貝
の貝殻利用であり、貝殻の遺物形成過程の複雑さを物語る遺物である。貝製品と考えられる貝殻が出土した場合には、その加工痕跡を慎重に観察し、製作技術を
検討し、遺物化過程を探ることが重要になってくる。この場合も現生の対象標本や民族誌や民俗例などの比較データが解釈に必要である。
さらに、いくら丈夫な貝殻であっても捕獲された貝殻が全て遺跡から検出されるわけではない。漆喰等の原料となる貝灰を製作する場合には、貝殻は高熱にさ
らされ著しく破損し、形状を失ってしまう。この場合、資料が出土しないため、現状では実証的解釈を行うのは困難であるが、一部でも貝殻組織が残存し、成長
線が観察できれば、同定できる可能性は残されている。
周辺遺跡での貝殻成長線分析の「比較データ」が得られたことで、貝類遺存体の形成過程が解釈できるようになった例もある。従来、縄文時代貝塚に捨てられ
た貝類は、捕獲されたほとんどであると考えがちであったが、貝塚以外に貝殻の廃棄場があったことが、明らかになったのである。アサリを主体とする西畑地点
と呼ばれる貝塚と、北側50m程の地点に隣接する製塩遺跡である西畑北地点が発掘された宮城県鳴瀬町里浜貝塚の場合、製塩遺跡からも多量のアサリとカキ類
が検出され、特にカキ類は製塩遺構周辺でより多く検出された。このことから、集落主体部に隣接する貝塚以外の地点にも、採集された貝殻が盛んに捨てられて
いたことがうかがわれた(小井川、加藤
1988)。これは食用貝類が全て貝塚に捨てられていたわけではないことを示すばかりでなく、人間が廃棄する場所と貝殻の種類を選択して、捨てていたこと
も示しており、食用貝類の遺物化の経路の複雑性を端的に表している。
富岡直人 2003 「貝殻
成長線分析」『環境考古学マニュアル』(同 成社):pp.237-250
環境考古学研究室: 岡山理科大学総合情報学部 生物地球システム学科 富岡研究室
トップページへ