| 食い痕があると、触角で丹念な探索を始め、 | 
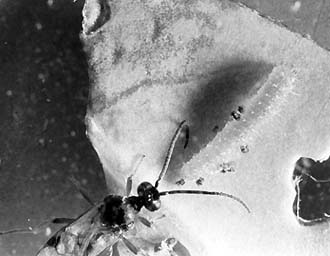 |
近くのアオムシを捜し当てる。 |
モンシロチョウに近縁のエゾスジグロシロチョウには寄生できるがその数は少ない。スジグロシロチョウには殆ど寄生できない。雌蜂は、モンシロチョウの食草上で若い齢のアオムシを捜し、産卵管を突き立て、体内に数十個の卵を産み込む。アオムシ体内で孵化したハチは内側から寄主を摂食し、本来ならアオムシが蛹化する頃に体表を食い破って中からコマユバチ幼虫が出てくる。幼虫はその場で直ちに繭を紡ぎ蛹化する。蛹の期間は一週間から十日ほどである。寄主はそのまま死んでしまう。
幼虫寄生蜂の中には、寄主の発育を抑えたり蛹化を妨げたりするものがいるが、アオムシコマユバチは寄主の発育を制御できない。齢の進んだ寄主に寄生すると、自らが発育を完了する前に寄主の蛹化時期が来てしまい、寄生に失敗することが多い。
|
アオムシコマユバチは身近な蝶である紋白蝶の幼虫(=青虫)に寄生するので、目にする機会が多い寄生蜂である。このハチの事を全般的に述べた一般向けの本はないが、部分的に触れた本が幾つかある。
「性の生物学1 ♂と♀のはなし」(1985) 越田豊 他、培風館。[57〜76頁] |
| 食い痕があると、触角で丹念な探索を始め、 |
|
近くのアオムシを捜し当てる。 |
ハチに寄生されるのは 3齢迄の若い幼虫が殆どだが、彼らは食草葉の裏にいる事が多い。親であるモンシロチョウが多くの卵を葉の裏面に産み付けるので、孵ったアオムシの多くが裏面にいるのは当然だ。しかし、表で孵ったアオムシも葉の下側が好きなのである。実は、葉裏にいるアオムシは葉表にいるアオムシに較べてアオムシコマユバチに寄生され難い。故に、アオムシは下側を好むのだと考えられる。そもそも、モンシロチョウが多くの卵を葉の裏面に産み付けるという行動自体が、孵化してくるアオムシを寄生蜂から守る行動ではないのか・・・。
Tagawa et al. (2008) Leaf surface preference in the cabbage worm, Pieris rapae crucivora, and parasitism by the gregarious parasitoid Cotesia glomerata. Entomol. Exp. Appl. 129: 37-43.
●アオムシのいない冬をどう過ごすのか?
モンシロチョウは寒くなると蛹の状態で休眠し越冬する。つまり、寄主であるアオムシは冬にはいなくなる。この蜂はどうやって冬越ししているのだろうか。
秋が深まり涼しい日が続くようになると健康なアオムシが休眠の準備に入る。寄生されていたアオムシからはハチ幼虫が脱出してくる。寄主から出て繭を紡いだ幼虫は、繭の中で直ぐに前蛹になる。暖かい頃だと一日もすれば蛹化するのだが、その頃に脱出して営繭したハチは前蛹の儘である。そして春になって暖かくなるまで蛹化しない。このハチは前蛹で休眠越冬している。
Ishii et al. (2000) Diapause in the braconid wasp, Cotesia glomerata (L.) II. Factors inducing and terminating diapause. Entomol. Science 3(1): 201-206.
Tagawa et al. (1984) Diapause in the braconid wasp, Apanteles glomeratus L. I. Evidence of diapause in overwintering prepupae. Appl. Entomol. Zool. 19(3): 396-399.
●雄と雌の出会いは?
多くの蜂は産雄性の性決定機構(haplo-diploid sex-determination system)を持っており、本種の未交尾雌も雄の子を産む事が出来る。この事が配偶行動や産卵行動の大本にある。雄の羽化は雌に比べて若干早い(protandry)。羽化直後の雄は、繭塊から発する雌の性フェロモンの匂いに惹きつけられて、しばらくその付近に留まる。遅れて羽化した雌を見つけると、後を追いかけ交尾する。うまくいかない場合は諦めてその場を去るが、結果として兄弟姉妹間での交配が高頻度で起こる事になる。しかし、同系交配率は6割程度であり、この種の多寄生蜂の同系交配率としてはそう高い方ではない。この事と関連して、母親の産む子ども達の性比が雌に偏っている。雌に偏った性比は、雄の交尾能力を反映したものでもある。
全ての雌が羽化直後に交尾するわけではなく、未交尾のまま羽化繭塊を離れる雌も多い。しかし野外雌の実際の交尾率は9割を超えているので、羽化直後に交尾できなかった蜂も、多くは其の後どこかで交尾の機会を持っている。
●寄生蜂に寄生する寄生蜂Tagawa (2002) Male insemination capability in the parasitoid wasp, Cotesia glomerata L. (Hymenoptera: Braconidae). Netherlands J. Zool. 52: 1-10.
雄は翅を震わせながら性フェロモン源に向かい、雌を見つけるといきなり後ろから乗りかかり交尾する。 雌は受身であり、積極的な行動を見せない。
Tagawa (2000) Sex allocation and clutch size in the gregarious larval endoparasitoid wasp, Cotesia glomerata. Entomol. Exp. Appl. 97: 193-202.
Tagawa (1992) Host discrimination by unmated individuals of the gregarious parasitoid wasp, Cotesia(=Apanteles) glomerata (Hymenoptera: Braconidae). Appl. Entomol. Zool. 27(2): 306-309.
Tagawa (1987) Post-mating changes in the oviposition tactics of the parasitic wasp, Apanteles glomeratus L. (Hymenoptera: Braconidae). Appl. Entomol. Zool. 22: 537-542.
Tagawa et al. (1987) Effects of mating on the oviposition behaviour of the parasitic wasp, Apanteles glomeratus L. (Hymenoptera: Braconidae). J. Ethol. 5: 37-41.
Tagawa et al. (1985) Influence of age on the mating behaviour of the braconid wasp, Apanteles glomeratus L. Appl. Entomol. Zool. 20: 227-230.
母蜂は産卵の前に寄主に麻酔をかける。その為、孵化幼虫は寄主を最大限に利用することが可能となる。 このハチは羽化直前にまで発育の進んだ寄主繭をも利用できるが、麻酔が無ければ古い繭は全く利用できない。
Tagawa et al. (2008) Development of the hyperparasitoid Eurytoma goidanichi Boucek (Hymenoptera: Eurytomidae) and the primary role of the mother wasp. Animal Biology 58: 113-121.
Tagawa et al. (2000) Longevity, egg load, and lack of host-quality preference in the hyperparasitoid Eurytoma goidanichi (Hymenoptera: Eurytomidae). Appl. Entomol. Zool. 35: 541-547.
Tagawa & Fukushima (1993) Effects of host age and cocoon position on attack rate by the hyperparasitoid, Eurytoma sp. [Hym.: Eurytomidae], on cocoons of the parasitoid, Cotesia(=Apanteles) glomerata [Hym. Braconidae]. Entomophaga 38: 69-77.

